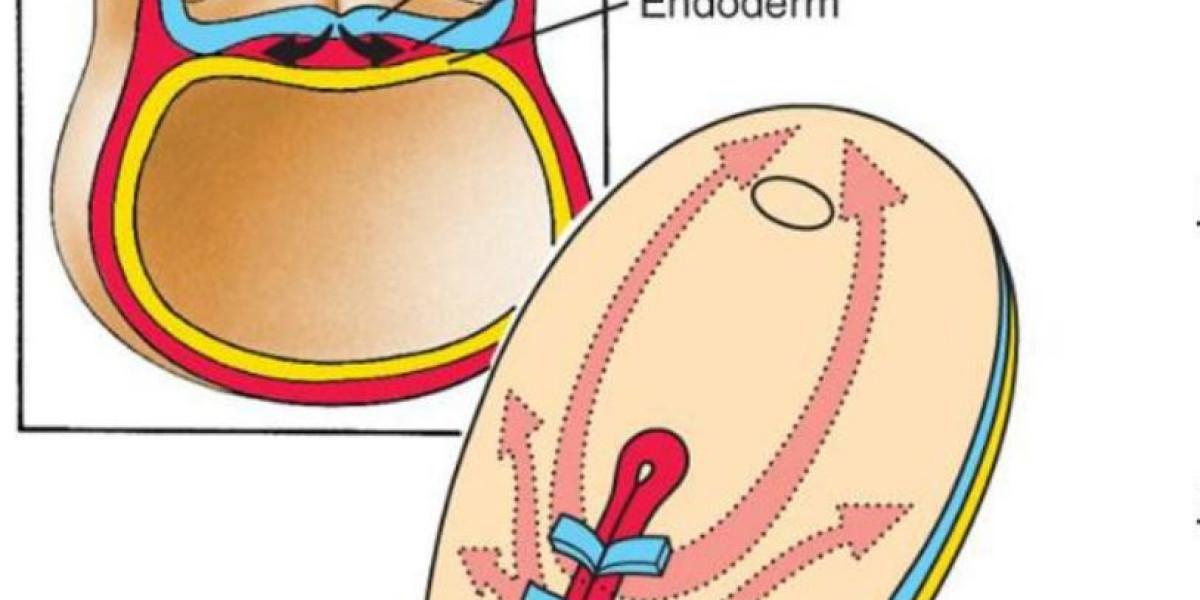
The gastrula refers to an animal embryo with a double germ layer or a triple germ layer, an important stage in the development of animal embryos. It is derived from the development of blastocysts. Part of the cells of the blastocyst migrates to the interior by various means to form the gastrula of the double or triple germ layer. Lower animals such as coelenterates consist of only two germ layers; many higher animals form mesoderm between the meat ectoderm. The cavity surrounded by endoderm is called the gut lumen. The migration of cells in gastrulation is called the gut effect. There are various ways, such as inset, outsourcing, stratification, migration, concentration, and extension. During this period, the nucleus began to play a leading role in the synthesis of new proteins, and the cells differentiated significantly, laying the foundation for tissue development and organogenesis.
Gastrulation
Gastrulation is a stage that most animal embryos experience. At this stage, only one layer of blastocysts recombines to form a gastrula containing three germ layers (ie, ectoderm, mesoderm, endoderm) cells. The gastrulation occurs after the cleavage. After the formation of the gastrula, the embryo enters the gastrula and begins the organogenesis process. The newly formed cells of the three germ layers will combine and develop into organs. The cells of each germ layer can develop into specific organs and tissues. The ectoderm develops into the epidermis, neural crest, and tissue that later develops into the nervous system. Mesoderm cells are located between ectodermal cells and endoderm cells and can develop into somite, muscles, and cartilage belonging to ribs and vertebrae. In addition, the mesoderm can also develop into the dermis, spinal cord, blood vessels and blood, bone, and connective tissue. Endoderm cells develop into the epithelium of the digestive system and respiratory system, such as the liver and pancreas. After the endocytic formation process, the cells of the body are organized into a group surrounded by cells associated with the epithelium. The molecular mechanisms of gastrulation and the time required are different in different organisms. However, gastrulation between different organisms still has something in common, such as: first, the topology of the embryo changes, from a single connected surface (like a spherical surface) to a non-single connected surface (like a ring) surface. Second, embryonic cells will differentiate into ectodermal cells, mesoderm cells, and endoderm cells (some lower organisms without mesodermal cells). Third, endoderm cells will have digestive function. In addition, although the specific patterns of animal gastrulation are very different, in general, during the process of gastrulation, the movement of cells can be summarized into five types: invagination, involution, ingression, delamination, and epiboly. In the preparation stage of gastrulation, the embryo forms a proximal-distal axis and an anterior-posterior axis, and asymmetry also occurs with the occurrence of these two axes. The formation of the embryonic egg cylinder marks the formation of the far and near axis: the proximal extraembryonic tissue forms the placenta-like tissue. Signal transduction pathways mediated by signaling molecules such as bone molding protein (BMP), fibroblast growth factor (FGF), Nodal, and Wnt are involved in this process. The visceral endoderm surrounds the epidermis. The distal visceral endoderm (DVE) migrates to the anterior part of the embryo, forming the anterior visceral endoderm (AVE), breaking the symmetry before and after. The above process is regulated by the Nodal signaling pathway. The original strip is produced in the initial stage of gastrulation. The original strip is located at the junction between the posterior extraembryonic tissue and the epidermis and the region where the internal migration occurs. The formation of the original streak is closely related to the Nodalsignaling pathway in cells in the coriolis region and the BMP4 signaling pathway activated by extra-embryonic tissues. Cer1 and Lefty1 can limit the formation of primitive streaks to specific regions by antagonizing the Nodal signaling pathway. The original strip area (after formation) will continue to grow distally. Many scientists are trying to compare the embryonic experiments within vitroexperiments to further study the process of gastrulation. Typically, researchers use 2D (traditional) or 3D (autologous cell culture methods)to culture embryonic stem cells (ESC) or artificially induced pluripotent stem cells (iPSC). Thein vitroculture method of the tissue culture method has lower cost, conforms to the 3R principle, and can accurately use the agonist/antagonist in a spatially and temporally specific manner compared to thein vivoexperimental method (in the intestine embryogenesis periodin vivo) It is difficult to do the third point. However,in vitroexperiments are somewhat different fromin vivoexperiments, so it is necessary to compare with the development of embryos in the body.
Germ layer formation
The germ layer, also known as the reproductive epithelium, is less used and is a group of cells when the animal embryo is formed. All animals have germ layers in which the vertebrate germ layer structure is particularly pronounced, while the sponge animal's germ layer is the simplest and usually produces two to three major tissue layers (sometimes referred to as primary germ layers). Radiation-symmetric animals (eg, coelenterates) have two germ layer structures, including endoderm and ectoderm; bilaterally symmetrical animals have three germ layer structures, more than radiation-symmetric animals, endodermal and external mesoderm between germ layers. All cells within the germ layer eventually develop into various tissues and organs of the animal. The inner, middle and outer blastoderm are named according to their position on the embryo at the end of the gastrula or at the beginning of the embryo. In fact, each layer of cells has its own characteristics in addition to the spatial position. For example, generally the endoderm cells are large, containing more yolk; ectodermal cells are small, and the division is fast. By means ofin vivostaining, the early distribution of the three germ layers can be determined in the gastrula or blastocyst stage, and the predicted fate of the embryo is depicted. This map essentially reflects that different germ layers may be associated with egg heterogeneity. It has long been known that many animals have regional differences in egg quality. For example, eggs have movements, and the distribution of macronuclei such as egg yolk particles, pigment particles, organelles, and nucleic acid proteins in various regions of the egg is unequal. When the cleavage divides the egg into many small pieces, the uneven oocyst is distributed into different lobes. As the gut movement progresses, the substances originally assigned to different lobes are distributed into different germ layers. These substances may affect further differentiation of each germ layer. For example, the lobes containing the yellow crescent region of the stalk will form muscle and interstitial cells in the future, and the lobes containing the gray crescent region will form the notochord and nerve tissue in the future, and the lobes containing the transparent material will form the epidermis in the future. The blastomeres of plant polar matter will form endoderm in the future. In general, the ectoderm forms the epidermis and nerve tissue. The endoderm forms the intestinal epithelium and the digestive gland epithelium, which forms bone, muscle, blood, lymph and other connective tissues. Others are derived from the mesoderm. The germ layer differentiation is basically along the route on the drawing. But there are exceptions: the sphincter of the eye iridescence does not come from the mesoderm, nor from the mesenchyme, but from a part of the retina, that is, from the ectoderm. The smooth muscle of the sweat gland is not from the mesoderm, but from the ectoderm; the mesenchyme itself is unclear, it may come from the ectoderm, or from the mesoderm, or even from the endoderm. As for the case of regeneration, dedifferentiation cells may change the direction of differentiation, and it is more difficult to distinguish the germ layer attached to it. The direction of germ layer development is not constant. In the echinodermal embryo, some mitotic balls are cut off, and the remaining blastomeres can compensate for differentiation into other types of tissues. Lithium ions convert the predetermined ectoderm of sea urchin into the structure of mesoderm, while thiocyanate or sulfate converts mesoderm and endoderm into the structure of the ectoderm. Similar changes can be made in the study of fish embryos and chicken embryos. The germ layer is only a preliminary division before differentiation. Some cytoplasmic components contained in the embryonic cells, the environment in which the cells are located, and the interaction between the cells play an important role in the direction of differentiation. The concept of germ layer is a pointer to the study of ontogeny and phylogeny. It helps to understand the intrinsic link between individual developmental changes and the development of various organs. For example, the retina of the eyeball is the structure in which the ectoderm is induced as well as the brain, and the olfactory sac and the ear sac are also structures in which the ectoderm is induced under other conditions. The pith of the adrenal gland comes from the ectoderm, while the cortex comes from the mesoderm, so their endocrine properties are very different. The concept of germ layer also helps to understand the evolutionary relationship between ontogeny and phylogeny. For example, a group of algae is a monolayer of cells, which can be equivalent to the blastocyst stage. Part of the intracellular and internal pleats of the blastocyst and coelenterates become a two-layer structure. The outer wall is the wall of the ectoderm, and the inside is the endoderm digestive tract.
Reference
- Solnicakrezel L, Sepich D S. Gastrulation: Making and Shaping Germ Layers.Annual Review of Cell Developmental Biology.2012, 28(28):687.
- Lyons D C, Perry K J, Henry J Q. Spiralian gastrulation: germ layer formation, morphogenesis, and fate of the blastopore in the slipper snail Crepidula fornicata.Evodevo.2015, 6(1):24.
- Nakanishi N, Sogabe S, Degnan B M. Evolutionary origin of gastrulation: insights from sponge development.Bmc Biology.2014, 12(1):26.
- Gaivo M M, Rambags B P, Stout T A. Gastrulation and the establishment of the three germ layers in the early horse conceptus.Theriogenology.2014, 82(2):354-365.
- Krieg M, Arboledaestudillo Y, Puech P H, et al. Tensile forces govern germ-layer organization in zebrafish.Nature Cell Biology.2008, 10(4):429-436.